
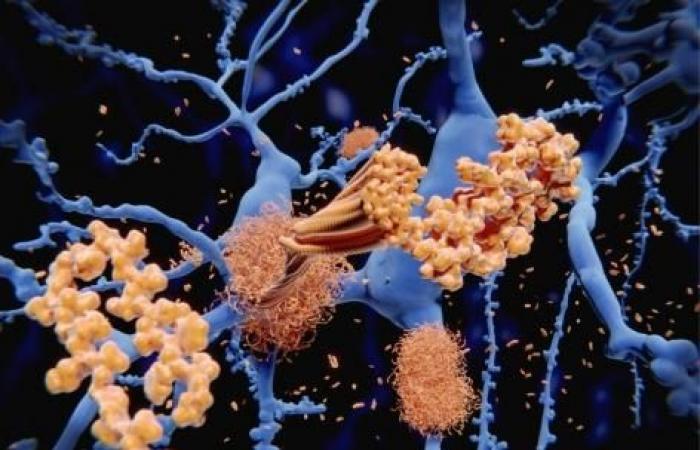
- تجمع پروتئین ها در بیماری های عصبی مانند آلزایمر
- تودههای پروتئینی بیماریزا در بیماری هایی مانند آلزایمر، در یک فرآیند وابسته به هستهزایی با زمان محدود تشکیل میشوند.
- پروتئین های تجمع یافته و ارتباطشان با وزیکول های EV
- اتصال EV و جذب متعاقب آن
- نقش گلیکوپروتئین های ویروسی در انتقال پلاک های پروتئینی
- گلیکوپروتئین های ویروسی، در طول عفونت حاد یا مزمن، می توانند توزیع پلاک های پروتئینی داخل بدن را تسهیل کنند.
- مکانیسم انتقال پلاک های پروتئینی آلزایمر
- روش انجام پژوهش
- نتایج مطالعه
- درباره نویسنده
تجمع پروتئین ها در بیماری های عصبی مانند آلزایمر
تجمع پروتئین ها، یک ویژگی پاتولوژیک بیماریهای عصبی (ND)، مانند بیماریهای آلزایمر (AD) و پارکینسون (PD) است. رسوب پروتئین مرتبط با بیماری معمولاً به صورت موضعی شروع می شود. متعاقباً به صورت کلیشه ای به سایر مناطق مغز گسترش می یابد. بیماری آلزایمر شایعترین بیماری تخریبکننده عصبی، با تجمع خارج سلولی آمیلوئید بتا و درون سلولی پروتئین تائو (Tau) است. تشکیل تودههای Tau نیز یکی از مشخصههای بیماری های تحلیل برنده عصبی است، در مجموع به عنوان تائوپاتی شناخته میشوند. انسفالوپاتی های قابل انتقال یا بیماری های پریون گروه خاصی از ND را تشکیل می دهند. این بیماری ها، بر انسان و سایر پستانداران تأثیر می گذارد. بیماری های پریون می توانند به صورت پراکنده و غیر ژنی رخ دهند. همچنین ممکن است در اثر جهش در ژن پروتئین پریون ایجاد شود. یا می تواند در اثر عفونت یا انتقال ایتروژنیک ایجاد شود.
تودههای پروتئینی بیماریزا در بیماری هایی مانند آلزایمر، در یک فرآیند وابسته به هستهزایی با زمان محدود تشکیل میشوند.
در این مرحله پروتئینهای مستعد تجمع الیگومرهایی تشکیل میدهند که به فیبریلهای مرتب و غنی از ورقه بتا تبدیل میشوند. انتقال بین سلولی تجمعات پروتئوپاتیک، به عنوان یک مکانیسم رایج ND در نظر گرفته می شود. مکانیسم دقیق انتقال این تجمعات بین سلولی و القاء پلاک های جدید نامشخص است. اما به نظر می رسد شامل آزادسازی پروتئین های آزاد، تماس مستقیم سلول به سلول توسط سیتونم ها باشد. همچنین تشکیل تونل های نانولوله ای (TNTs) یا EV نیز در این انتقال نقش دارد. EV ها وزیکول های ارتباطی نانومتری هستند که تحت شرایط فیزیولوژیکی و آسیب شناسی ترشح می شوند. EV در منشأ سلولی خود متفاوت است. برخی از آنها مستقیماً از سطح سلول جوانه می زنند (میکرووزیکول). برخی دیگر هنگامی ترشح می شوند که اجسام چند وزیکولی با منشا آندوزومی با غشای پلاسما ترکیب می شوند. در نتیجه اگزوزوم ها را به فضای خارج سلولی آزاد می کنند.
پروتئین های تجمع یافته و ارتباطشان با وزیکول های EV
وزیکول های ترشح شده همپوشانی قابل توجهی با میکرووزیکول ها و اگزوزوم ها دارند. این همپوشانی شامل اندازه، نشانگرهای سطحی و عملکرد، در نتیجه به عنوان EV شناخته می شوند. پروتئینهای محلول و انباشته با انواع بیماری های عصبی مانند آلزایمر مرتبط هستند. این ها توسط نورونها و سایر سلولها یا به عنوان پروتئینهای آزاد یا در ارتباط با EV ترشح میشوند. عملکرد این پلاک های پروتئینی در القاء بیماری ها در شرایط آزمایشگاهی به خوبی بررسی نشده است.
فقط بخش کوچکی از پروتئین های محلول یا تجمع یافته آزاد شده با EV مرتبط هستند. در حالی که اکثریت قریب به اتفاق آزادانه ترشح می شوند. به عنوان مثال، کمتر از 1٪ از کل آمیلوئید بتا ترشح شده با EV مرتبط است. تنها 3٪ از کل آلفا-سینوکلئین (α-synuclein) ترشح شده در وزیکول EV یافت شد. برای آزادسازی محموله سیتوزولی، EV معمولا با غشای سلولی ادغام می شود.
اتصال EV و جذب متعاقب آن
اینها فرآیندهای انتخابی هستند که نیاز به فعل و انفعالات غشایی خاص دارند. در نتیجه، فعل و انفعالات گیرنده-لیگاند بین EV و سلول های گیرنده، احتمالاً انتشار محموله پروتئوپاتیک تعدیل می شود. اینتگرینها و پروتئوگلیکانهایی شناسایی شده موجب اتصال EV به سلولهای هدف میشوند. اکثر جفتهای گیرنده-لیگاند که زیربنای این تعاملات هدفمند هستند، تاکنون ناشناخته هستند.
احتمالا فعالیت ضعیف القای تجمع برخی از EV حاوی پلاک ها می تواند به دلیل فقدان لیگاندهای خاص مورد نیاز برای فعل و انفعالات گیرنده میزبان و/یا همجوشی باشد. تماس و همجوشی غشاء می تواند توسط گلیکوپروتئین های ویروسی تسهیل شود. این گلیکوپروتئین ها به ویروس ها اجازه می دهند به سلول های هدف خود بچسبند و به آنها نفوذ کنند. تماس با گیرنده ها منجر به تغییر شکل در گلیکوپروتئین های ویروسی می شود. در نتیجه دو غشا را در مجاورت خود قرار می دهد و ادغام دو لایه را تقویت می کند.
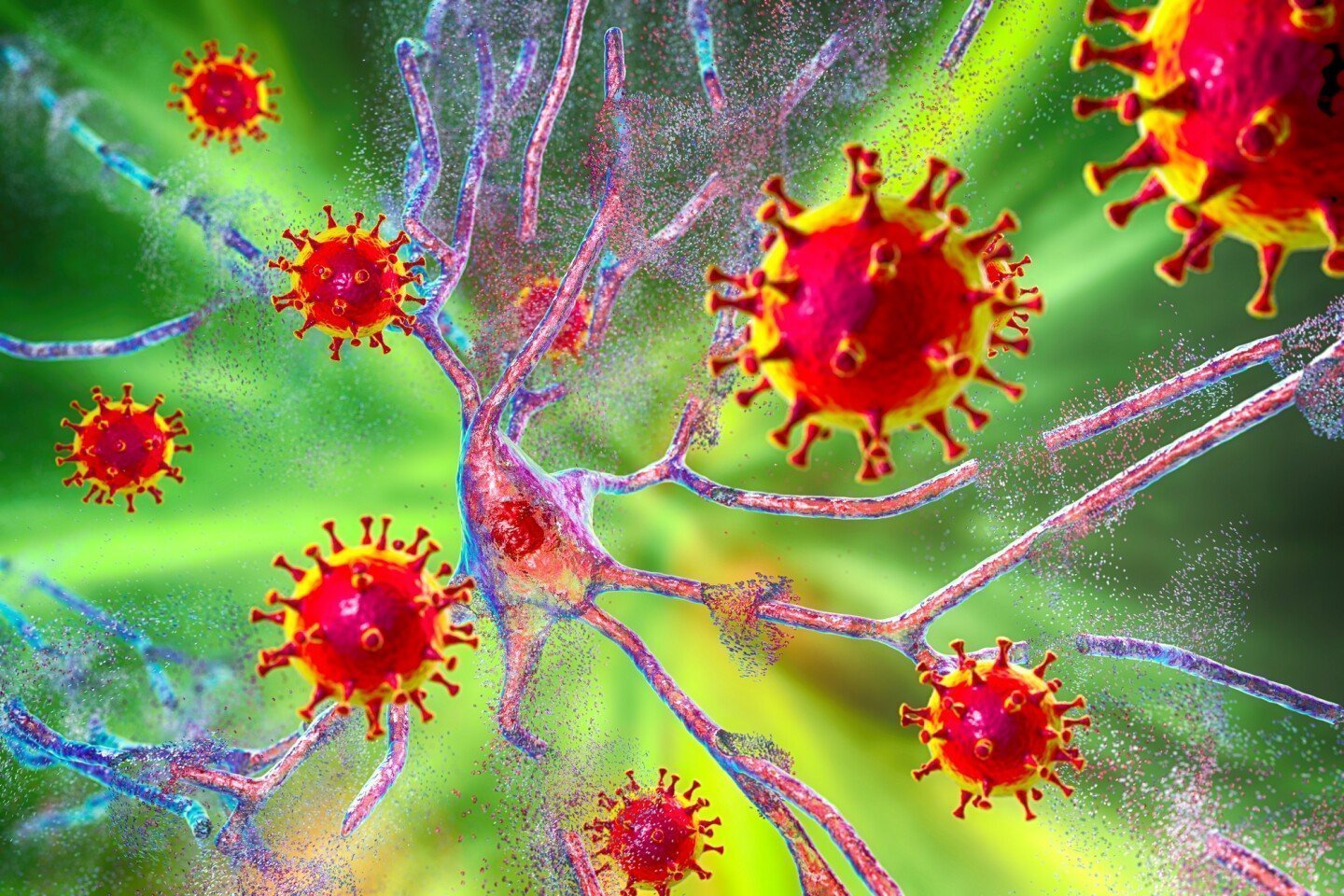
نقش گلیکوپروتئین های ویروسی در انتقال پلاک های پروتئینی
گلیکوپروتئین VSV-G ویروس استوماتیت تاولی تنها پروتئین سطحی این ویروس متعلق به جنس رابدویروس است. واسطه اتصال به گیرنده LDL یا اعضای خانواده آن است. VSV-G خارج از رحم در سطح سلول یافت می شود. علاوه براین، EV را در غیاب سایر اجزای ویروسی می پوشاند. اخیراً نشان داده شده است که VSV-G تحویل محموله با واسطه EV به سلول های گیرنده را افزایش می دهد. ما در اینجا VSV-G را در مدلهای سلولی که پلاک های پروتئینی مختلف را منتشر میکنند بیان کردیم.
همچنین، بیان VSV-G باعث ترشح EV با پوشش VSV-G با افزایش ظرفیت القاء پلاک در سلولهای گیرنده می شود. فعل و انفعالات بین پروتئین اسپایک کرونا ویروس-2 و گیرنده آن در گسترش پریونهای سیتوزولی و تائو کمک کرد. بنابراین، انتقال پلاک پروتئوپاتیک بین سلولی کارآمد را می توان به شدت توسط فعل و انفعالات گیرنده-لیگاند کنترل کرد.
گلیکوپروتئین های ویروسی، در طول عفونت حاد یا مزمن، می توانند توزیع پلاک های پروتئینی داخل بدن را تسهیل کنند.
برخی از بیماری های ویروسی ممکن است به تخریب سلول های عصبی کمک کنند. محققان DZNE این را در مجله علمی “Nature Communications” گزارش کردند. ارزیابی آنها بر اساس آزمایشهای آزمایشگاهی است. آنها توانستند نشان دهند که مولکولهای ویروسی خاص، گسترش بین سلولی تودههای پروتئین را تسهیل میکنند. این توده های پروتئینی نشانه بیماریهای مغزی مانند آلزایمر هستند.
یافتهها ممکن است سرنخهایی را ارائه دهد که چگونه عفونتهای ویروسی حاد یا مزمن میتوانند به تخریب عصبی کمک کنند. تودههای پروتئینهای تاخورده نادرست توانایی انتقال از یک سلول به سلول دیگر را دارند. در آنجا شکل غیرطبیعی خود را به پروتئینهایی از همان نوع منتقل میکنند. در نتیجه، این بیماری در سراسر مغز گسترش می یابد.
پدیده مشابهی برای آلزایمر و بیماری پارکینسون مورد بحث قرار گرفته است. انتقال تودهها میتواند شامل تماس مستقیم سلول به سلول، انتشار تودههای «برهنه» در فضای خارج سلولی یا بستهبندی وزیکولی باشد.
مکانیسم انتقال پلاک های پروتئینی آلزایمر
رهبر گروه تحقیقاتی می گوید: “مکانیسم های دقیق انتقال ناشناخته است.” وزیکول ها حبابهای کوچکی هستند که توسط یک پوشش لیپیدی احاطه شدهاند و برای ارتباط بین سلولها ترشح میشوند. با این حال، چند پیشنهاد مطرح شده است. تبادل از طریق تماس مستقیم سلولی و هم از طریق وزیکول ها به تعاملات لیگاند-گیرنده بستگی دارد. در هر دو حالت، غشاها نیاز به تماس و فیوز شدن دارند. هنگامی که لیگاندهایی وجود دارند که به گیرنده های سطح سلول متصل می شوند. در نتیجه باعث ادغام دو غشا می شوند، این امر آسان تر می شود”.
انتقال بین سلولی پریون ها یا توده های پروتئین های تائو بررسی شد. زیرا آنها به شکل مشابه در بیماری آلزایمر و سایر “تائوپاتی ها” رخ می دهند.محققان با تقلید از آنچه در نتیجه عفونت ویروسی اتفاق میافتد، سلولها را وادار به تولید پروتئینهای ویروسی کردند. اینها واسطه اتصال سلول هدف و همجوشی غشاء است.
روش انجام پژوهش
دو پروتئین بهعنوان نمونههای اصلی انتخاب شدند. پروتئین اسپایک کرونا و گلیکوپروتئین VSV-G ویروس استوماتیت تاولی، که در پاتوژن آلوده کننده گاو و سایر حیوانات نقش دارد. علاوه بر این، سلولها گیرندههایی را برای این پروتئینهای ویروسی، یعنی خانواده گیرندههای LDL، بیان کردند. اینها به عنوان پورت اتصال برای VSV-G و ACE2 انسانی، گیرنده پروتئین اسپایک عمل میکنند.
مشاهده شد که پروتئینهای ویروسی هم در غشای سلولی و هم در وزیکولهای خارج سلولی ترکیب میشوند. وجود آنها باعث افزایش تجمع پروتئین بین سلول ها به دو طریق می شود. از طریق تماس مستقیم با سلول و از طریق وزیکول های خارج سلولی. لیگاندهای ویروسی واسطه انتقال موثر پلاک ها به سلولهای گیرنده شدند. همچنین آنها ایجاد پلاک های جدید را القا کردند. ووربرگ میگوید لیگاندها مانند کلیدهایی عمل میکنند که قفل سلولهای گیرنده را باز میکنند بنابراین مخفیانه وارد محموله خطرناک میشوند.
نتایج مطالعه
دادههای ما نشان میدهد که فعل و انفعالات لیگاند-گیرنده ویروسی میتواند بر انتقال پروتئینهای پاتولوژیک تأثیر بگذارد. این یک یافته جدید است.
اثرات بالقوه بر تخریب عصبی مغز بیماران مبتلا به بیماریهای عصبی گاهی اوقات حاوی ویروسهای خاصی است. آنها مشکوک به ایجاد التهاب یا اثر سمی هستند، بنابراین تخریب عصبی را تسریع می کنند. با این حال، پروتئینهای ویروسی نیز میتوانند متفاوت عمل کنند. آنها میتوانند گسترش بین سلولی تودههای پروتئینی را که در حال حاضر در بیماریهای تخریبکننده عصبی مانند آلزایمر ادامه دارند، افزایش دهند.
” تأثیر عفونتهای ویروسی بر بیماریهای نورودژنراتیو نیازمند بررسی عمیق است”.
برخی از عفونت های ویروسی می توانند گسترش بین سلولی توده های پروتئینی مرتبط با اختلالات عصبی را افزایش دهند. در نتیجه خطر ابتلا به آلزایمر، پارکینسون و سایر بیماری های عصبی را افزایش دهند. تبادل محموله سلولی با تماس مستقیم سلولی یا از طریق EV بستگی به برهمکنش گیرنده-لیگاند دارد.
منبع: https://neurosciencenews.com/viral-infections-neurodegeneration-19497/

درباره نویسنده
خجسته رحیمی جابری، پژوهشگر دکترای تخصصی علوم اعصاب دانشگاه علوم پزشکی شیراز و نویسنده وب سایت انستیتو سلامت مغز دانا. زمینه کاری تخصصی ایشان مشکلات حافظه و آلزایمر است وی بیش از 20 مقاله معتبر بین المللی در این زمینه به چاپ رسانده است. می توانید پژوهش های او را در اینجا (کلیک کنید) دنبال کنید.